
Foi em 1924 que Hans Berger obteve o primeiro traçado eletrencefalográfico humano. Utilizando eletrodos de agulha introduzidos sob o couro cabeludo e um eletrocardiógrafo, Berger descobriu nas áreas occipitais as oscilações de potencial que denominou "ondas alfa". Este pesquisador estudou durante vários anos o eletrencefalograma (EEG) humano, publicando seus trabalhos entre 1929 e 1939. Adrian, em 1934, empreendeu também a análise das ondas cerebrais e de seus trabalhos resultou o interesse que durante e após a segunda guerra mundial levou à aplicação da eletrencefalografia à prática médica e à investigação neurofisiológica.
Embora tenha sido Berger o primeiro a descrever o EEG humano, coube a Caton, fisiologista inglês, descobrir em 1875 que era possível registrar variações de potencial no córtex cerebral. A atividade eletrofisiológica de nervos estava então sendo amplamente estudada na Alemanha e Caton buscava no cérebro fenômenos semelhantes ao potencial de ação, quando verificou que, registrando a diferença de potencial entre dois pontos do córtex cerebral exposto de coelhos, ocorriam espontaneamente oscilações elétricas, as quais podiam ser modificadas por estimulação sensorial. Caton provou que tais potenciais eram de natureza biológica, pois podiam ser abolidos por anóxia, anestesia e morte3.
Se o registro da atividade elétrica cerebral (AEC) é hoje tecnicamente muito fácil, analisar sua significação e seus mecanismos envolve problemas e metodologia muito complexos. Quando Adrian estudou o EEG humano preocupou-se com a origem das ondas, porém pouco mais pôde fazer que concluir pela existência de algum marca-passo subcortical que controla, pelo menos, a sincronia do ritmo alfa. Um exame mais aprofundado da questão revela que a análise objetiva da AEC abrange um conjunto de problemas neuro-fisiológicos muito complicados, dos quais se salientam os seguintes: 1) origem dos potenciais; 2) modificação ou modulação da AEC por estímulos sensoriais, alterações do estado de vigília, de sono, de atenção; 3) significação fisiológica desses fenômenos eletrofisiológicos.
Uma exposição razoavelmente completa sobre todos estes aspectos seria sobremodo longa e talvez pecasse por excesso de informações de difícil assimilação. Limitar-nos-emos, por isso, a discutir alguns aspectos fundamentais dos mecanismos de gênese da AEC.
Eletrofisiologia da membrana neuronal — A membrana celular é constituída por uma dupla camada de lipóides (cujas extremidades polares estão voltadas para dentro e para fora) e uma de proteínas de cada lado, atingindo 80 angstroms de espessura. A combinação de lipóides de cada tipo de membrana parece ser determinada geneticamente; no axônio a parte não polar é constituída principalmente de ácidos graxos com 16 a 20 átomos de carbono e a extremidade polar dos diversos lipóides varia tanto em tamanho como em configuração e carga elétrica. A microscopia eletrônica revelou que, externamente ã membrana celular, os neurônios dispõem de uma outra camada, denominada membrana exterior, cuja opacidade aos elétrons é muito débil, o que dificulta sua identificação. A espessura da membrana exterior é de aproximadamente 75 angstroms e em sua constituição entram predominantemente glicolípides e glicoproteinas.
Devido à diferente composição iônica dos líquidos intra e extracelular estabelece-se, através da membrana celular, uma diferença de potencial que pode ser calculada desde que se conheçam as concentrações de certos íons (potássio, sódio e cloro), sobretudo o potário (K+). A concentração de K + no líquido intracelular é, nos mamíferos, cerca de 40 vezes maior que no extracelular. Esse gradiente de concentração acarreta forte tendência de os íons K + passarem para o lado de fora da membrana. Em virtude de sua mobilidade ser muito maior que a dos aníons a ele ligados no interior da células, o K+ migra para o lado externo, deixando atrás de si os aníons que não conseguem atravessar a membrana por serem de grande porte, se comparados com o K+ . Nessas condições estabelece-se separação de cargas, com a frente positiva no lado externo, determinada pelo K+, em contraposição à negativa interna, deteiminada por íons aspártico, fumárico, glutâmico e outros Isso leva a uma diferença de potencial elétrico através da membrana, sendo a face externa positiva em relação ã interna. A saída de K+ não cresce indefinidamente, porquanto a elevação da diferença de potencial logo começa a opor-se eletrostàticamente à saída de K+ . Como conseqüência disso, em dado momento a força (devida à diferença de concentração) que faz os íons K+ sairem da célula é contrabalançada pela diferença de potencial gerada (e que atrai K+ para dentro). Nesse instante ocorre equilíbrio dinâmico, de tal forma que o número de íons K+ que saem é igual aos que entram na célula. O potencial elétrico que equilibra a força originada pela diferença de concentração de K denomina-se potencial de equilíbrio e pode ser calculado pela equação de Nernst:

na qual E = potencial de equilíbrio para o K ; R = constante dos gases perfeitos; T = temperatura absoluta; F = constante de Faraday; Ki = concentração interna de K+ ; Ke. = concentração externa de K+.
A equação de Nernst estabelece que o potencial de equilíbrio do K + é proporcional ao logaritmo da relação entre as concentrações interna e externa. Esse potencial é aproximadamente igual ao potencial de membrana, medido por intermédio de um eletrodo situado no interior da célula e outro no meio extracelular, quando a célula está em repouso funcional.
Enquanto a concentração de K+ é maior no meio intracelular» a concentração de sódio (Na+ ) é maior no extracelular, onde é mantida em níveis elevados porque a membrana tem um dispositivo metabólico que expulsa ativamente íons Na+ no estado de repouso. Se o neurônio é estimulado a estrutura da membrana se deforma e permite a entrada abrupta de Na. Duas situações podem então ocorrer: se o estimulo é pouco intenso (sub-limiar), a entrada de Na+ reduz o potencial de membrana; se o estímulo é intenso (limiar ou supralimiar), a entrada de Na+ é maciça e promove a inversão do potencial de membrana, pois a separação dos íons de Na+ causa uma diferença de potencial como para o K+ , agora porém com a frente positiva voltada para dentro. No primeiro caso é possível registrar a ativação como pequenas reduções do potencial de membrana (despolarizações, potenciais locais, graduados ou eletrotônicos). Tais potenciais se propagam a apenas poucos milímetros ou frações de milímetro do ponto estimulado. Aumentando-se ligeiramente a intensidade do estimulo a despolarização aumenta e facilita a ativação completa da membrana por outros estímulos sub-limiares. Quando, porém, o estímulo é forte, a despolarização é grande e atinge um valor, denominado limiar, que desencadeia um aumento da permeabilidade da membrana ao sódio, levando à inversão do potencial. Ao contrário do que acontece com os potenciais locais, a inversão é um potencial tudo-ou-nada, uma vez que, depois de atingido o limiar, a intensidade do estimulo não o modifica, desde que seja suficiente para provocar um valor igual ou muito maior do que o limiar. Outra diferença importante entre os dois tipos de resposta é que a inversão se propaga ao longo do axônio com amplitude mais ou menos constante, regenerando-se o processo de um ponto a outro até chegar à extremidade. Nas células nervosas essa onda de inversão propagada denomina-se impulso nervoso c pode ser registrada por meio de eletrodos colocados no lado externo ou um no lado externo e outro no interno. O potencial assim registrado denomina-se potencial de ação, que é a manifestação eletrofisiológica do impulso nervoso.
A velocidade de propagação do potencial de ação varia de acordo com o diâmetro da fibra nervosa e com a natureza do tecido. É maior nas fibras mais grossas (fibras motoras, proprioceptivas, fibras do trato piramidal e do trato vestibulospinal), nas quais pode chegar a 120 metros/segundo, do que nas finas (fibras aferentes cutâneas térmicas e dolorosas). É maior nas fibras nervosas grossas do que nas fibras musculares. Nas fibras finas e curtas do sistema nervoso central é ainda incerto que realmente ocorram impulsos nervosos; é provável que a transmissão seja em grande parte de natureza eletrotônica. De qualquer forma essa velocidade é pequena (frações de milímetro por segundo), mas as distâncias a percorrer são, também, extremamente curtas.
Após a inversão da polarização há inversão do processo iônico e refaz-se a diferença de potencial de repouso da membrana, ficando ela pronta a receber novo estímulo. Fisiologicamente isso podo ocorrer centenas de vezes por segundo. As fibras do trato piramidal, por exemplo, podem conduzir impulsos com a freqüência de 800-1000 por segundo.
A transmissão de informações de um elemento para outro, pelo menos nos vertebrados, se faz por intermédio de um dispositivo anatômico altamente diferenciado, a sinapse. Em invertebrados a transmissão pode ser feita, em muitos sistemas, de uma fibra a outra diretamente, no ponto de justaposição, que freqüentemente é uma decussação. Na maioria dos sistemas neurais, porém, as sinapses são constituídas de elementos especializados que permitem estabelecer as condições de transmissão de informações. A terminação axônica se resolve em múltiplos botões terminais, que perdem a camada de mielina (se a fibra é mielíniea) e se apõem à membrana da célula seguinte. A membrana da terminação denomina-se pré-sináptica e a da célula seguinte chama-se pós-sináptica. Entre ambas existe um delgadíssimo espaço, cheio de líquido intersticial, denominado fenda sináptica. No interior da terminação encontram-se numerosas vesículas que contêm moléculas dissolvidas de substâncias responsáveis pela ativação da membrana pós-sináptica, de acordo com o seguinte mecanismo: em condições de repouso funcional da sinapse algumas vesículas saem da terminação ao acaso e libertam na fenda sináptica o seu conteúdo (chamado genericamente mediador químico); a acetilcolina e a noradrenalina são os únicos até o momento indubitavelmente identificados como mediadores. Entrando em contato com a membrana pós-sináptica o mediador combina-se com certos radicais e da combinação resulta a formação de complexos moleculares, que deformam a membrana permeável ao sódio extracelular, causando pequena despolarização, insuficiente para causar ativação da célula; esse processo repete-se muitas vezes no tempo e no espaço e é detectável eletrofisiològicamente sob a forma de pequenos potenciais (minipotenciais) eletrotônicos. Quando chega um impulso à terminação a libertação do mediador é subitamente multiplicada centenas de vezes, de forma que os minipotenciais se somam e podem atingir o limiar do neurônio. Se isso acontece, um potencial de ação nasce no segmento inicial do axónio e se propaga a toda a célula.
Quando há necessidade de inibir um neurônio o complexo molecular formado com o mediador na membrana pós-sináptica causa aumento da permeabilidade ao K+ ou ao Cl e isso, como seria de esperar, acarreta aumento do potencial de membrana localmente, havendo hiper polarização em vez de despolarização. Os potenciais eletrotônicos de despolarização, implicados na excitação celular, denominam-se potenciais excitatórios pós-sinápticos (PEPS); os envolvidos na inibição são os potenciais inibitórios pós-sinápticos (PIPS). Em suma, pois, os processos de excitação são precedidos de PEPS e os de inibição, de PIPS9,10.
MECANISMO E SIGNIFICAÇÃO FUNCIONAL DAS ONDAS CEREBRAIS
As ondas cerebrais representam a resultante de vários tipos de fenômenos eletrofisiológicos que, por sua vez, refletem numerosas operações neurais. Em que pese a vultosa coleção de dados sobre essas operações e aquelas manifestações ainda não é possível estabelecer se as ondas cerebrais, como fenômenos eletrofisiológicos, desempenham alguma função específica ou se constituem apenas um epifenómeno da atividade neural.
Uma das possíveis funções da AEC é a modificação da excitabilidade dos neurônios pelas correntes resultantes da atividade do conjunto. Há muitas evidências experimentais de que isso ocorre, especialmente no que diz respeito a potenciais muito lentos; mesmo para o ritmo alfa parece certo que a excitabilidade cortical e o tempo de reação a um estímulo aferente sofrem modificações de acordo com a fase da onda alfa que esteja ocorrendo no momento em que se aplique o estímulo. O problema, contudo, está ainda em discussão, porquanto há experiências discrepantes. Ainda que se consiga algum dia demonstrar que as ondas cerebrais atuam sobre os elementos neurais de forma definida, não será fácil saber se suas características (amplitude, freqüência, sentido das correntes, relação de fase entre potenciais de áreas vizinhas) são organizadas deterministicamente para provocarem os efeitos que causam ou se causam tais efeitos secundariamente por terem aquelas características.
Esse problema está longe de ser solucionado. O mecanismo das oscilações, porém, já se esboça como uma associação de fenômenos compreensíveis. A investigação desses mecanismos é sumamente difícil, sobretudo por dificuldades de natureza técnica. É impossível conseguir dados simultâneos de mais do que uns poucos neurônios em uma área cortical qualquer. Considerando-se que a densidade de neurônios pode chegar a milhares por milímetro cúbico pode-se ter idéia da extrema limitação técnica com que nos debatemos. Uma solução parcial dessa dificuldade é analisar as manifestações eletrofisiológicas de fenômenos corticais transitórios e relativamente localizados, os quais discutiremos mais adiante.
Os fenômenos eletrofisiológicos que contribuem para as ondas cerebrais são, ao que tudo indica, tanto as alterações parciais do potencial de membrana dos neurônios corticais (PEPS e PIPS) como suas descargas (potenciais de ação) e outros potenciais muito lentos que podem decorrer do conjunto de ação de agregados neuronais amplos1. Os neurônios corticais, agrupados segundo circuitos muito complexos, interagem uns com os outros. Cada um dos neurônios de um circuito deve entrar em ação nos momentos precisos; há instantes em que determinados neurônios devem estar inibidos, para não inteferirem com o funcionamento de outros; nesses momentos eles recebem impulsos inibidores (hiperpolarizantes) a fim de se tornarem inertes ante os impulsos que lhes chegam de diversas fontes. Por outro lado, alguns neurônios são ativados pelos impulsos de outros, que em certas condições podem ser os mesmos que inibem os primeiros acima referidos. Nem sempre esses neurônios realmente descarregam impulsos, restringindo-se todo o processo apenas a PEPS. O resultado é que as correntes excitadoras e inibidoras, que têm sentidos opostos em relação aos pontos celulares inibidos ou estimulados, se compõem e são registradas como oscilações regulares ou irregulares de potencial. Teoricamente, portanto, é de se esperar que, com dispositivos especiais, se possam surpreender os potenciais elementares cuja composição redunda nas oscilações eletrencefalográficas. De fato, o registro do EEG com filtros que permitem a passagem de faixas estreitas de freqüência demonstra a existência de freqüências de 20, 100, 120, 200 e até 500 Hz nos mesmos pontos em que o registro usual evidencia ritmos de baixa freqüência.
Vejamos agora, sucintamente, quais são as provas experimentais de que as ondas cerebrais representam a composição de potenciais eletrotônicos e potenciais de ação de neurônios corticais.
1. Potenciais locais de superfície — A excitação direta do córtex por estímulos elétricos causa resposta bem típica na superfície, constituída de um potencial negativo de breve duração, seguido de outro positivo de baixa voltagem. Se o estímulo é intenso obtém-se um terceiro potencial, agora negativo, gerando um complexo muito semelhante a certas manifestações de focos epileptógenos. O primeiro potencial se deve à ocorrência de PEPS nos 400-500 micra mais superficiais do córtex; o potencial positivo deve-se à ocorrência de PIPS, aparentemente nas mesmas camadas (Fig. 1). Um eletrodo colocado mais profundamente não registra alterações da AEC ao estímulo fraco e efêmero. Esse fato, além de outros que não cabe aqui referir, introduz séria complicação na interpretação dos potenciais corticais. De há muito se considera que o cérebro se comporta elètricamente como um condutor em volume e que, portanto, as alterações de potencial em certa área podem ser registradas em outras, sempre com modificações de forma, amplitude e fase que dependem da distribuição física das correntes num condutor tridimensional homogêneo. No entanto, esse conceito só tem valor relativo, uma vez que a modificação de posição dos eletrodos não só modifica sua posição em relação às linhas de corrente mas também os põe em contato com as proximidades de outras fontes de potencial. O que é válido para um nervo submetido a estímulos padronizados não pode ser válido para estruturas em que a ativação de um elemento acarreta modificações funcionais em outros, de forma conhecida ou desconhecida.

Grundfest e col. 11 desenvolveram ampla análise dos componentes de potenciais elementares corticais, utilizando drogas que seletivamente alteram um ou outro componente. Esses autores verificaram, por exemplo, que a aplicação tópica de GABA (ácido gama-aminobutírico) no ponto em que se registra um potencial local, obtido por estímulo elétrico direto no córtex cerebral, abole totalmente a resposta negativa, que resulta de PEPS, e evidencia uma positiva, resultante de PIPS. A mesma experinécia, feita no córtex cerebelar, leva à abolição do componente negativo mas não evidencia o positivo (Fig. 2). Essas experiências demonstram não só que um mesmo tipo de potencial, nc caso a resposta local cerebral e a cerebelar, embora semelhantes podem envolver componentes diversos e, portanto, informações distintas, mas também que mesmo um potencial simples pode conter elementos inaparentes porque são mascarados.

O segundo componente positivo da resposta local era tido como ocasionado pela ativação maciça de interneurônios corticais. Mas Yamamoto e Kawai22 verificaram, recentemente, que mesmo esse componente é de origem superficial e, portanto, depende dos dendritos apicais. Esses autores conseguiram isolar e manter viva apenas a carnada molecular de blocos de córtex cerebral, contida em uma lâmina de 200 micra de espessura, e registraram típicos potenciais locais em resposta a estímulos breves. Como a camada molecular contém as ramificações apicais dos dendritos das células piramidais essas experiências confirmaram que respostas locais têm origem nessas fibras e independem das conexões intercorticais; o segundo componente positivo só pode originar-se nas fibras, visto que nessa camada praticamente não há corpos celulares.
2. Potenciais evocados, aumentantes e de recrutamento — A estimulação elétrica ou fisiológica de um órgão sensorial provoca, desde que o estimulo seja brusco e efêmero, um potencial bem característico na área cortical sensorial correspondente. Após curta latência ocorre um potencial positivo breve, seguido de outro negativo de duração maior. A esse complexo seguem-se pequenas oscilações cujas características, como no caso do primeiro e do segundo potenciais, dependem da região em que se localiza o eletrodo receptor ativo.
A excitação iterativa dos núcleos relés talâmicos (VPL, VPM, VL) causa, nas áreas corticais somestésica e motora, potenciais semelhantes aos potenciais evocados porém de amplitudes crescentes (daí o nome de potenciais aumentantes que se lhes dá).
A excitação iterativa a baixa freqüência dos núcleos do sistema reticular talâmico (núcleos intralaminares e da linha média) provoca o aparecimento, em certas áreas corticais, de potenciais de recrutamento, assim chamados porque crescem progressivamente até atingirem certa amplitude e, depois, oscilam periodicamente de voltagem enquanto dura a estimulação. Ao contrário do que ocorre com os potenciais evocados, porém, o primeiro potencial é negativo e o segundo, que surge em geral depois do terceiro ou quarto potencial, é positivo. Essas respostas são idênticas aos potenciais elementares que constituem os fusos tão característicos das primeiras fases do sono.
Por serem potenciais transientes que podem ser seguramente controlados e cronàdos as três modalidades eletrofisiológicas acima descritas foram objeto de múltiplos estudos no sentido de se correlacionarem seus componentes com fenômenos elementares dos neurônios corticais e talâmicos2,8,12,I5,20. Esses estudos demonstraram claramente que existe relação entre as ondas registradas no córtex e as modificações do potencial de membrana de neurônios corticais. Creutzfeldt e col.6, registrando simultaneamente o eletrocorticograma e os potenciais intracelulares de neurônios do córtex visual do coelho, verificaram que as ondas corticais positivas correspondem a despolarizações celulares (PEPS) e as negativas a hiperpolarizacões (PIPS); o mesmo acontece no córtex auditivo mas na área motora ocorre o oposto, possivelmente devido à diversidade dos padrões de conexões ou do tipo de neurônio estudado. No que respeita a potenciais evocados e potenciais aumentantes Uchida20 descreveu, no gato, o mesmo tipo de correlação.
Creutzfeldt e col.8 desenvolveram recentemente engenhosa técnica de análise da correlação entre ondas e potenciais celulares e com ela puderam reforçar suas observações anteriores, ao mesmo tempo em que deixaram claro que essa correlação é mais complexa do que aparenta. Eles extraíram de traçados simultâneos, feitos com macro-elétrodos de superfície e micro-elétrodos intracelulares, potenciais de EEG semelhantes e os superpuseram, de modo a obterem ondas relativamente homogêneas em forma e amplitude. Fazendo o mesmo com os traçados simultâneos obtidos com micro-elétrodos e depois comparando-os, puderam evidenciar claramente que existe correlação entre as duas manifestações eletrofisiológicas. As ondas puramente negativas são quase simétricas em relação ao pico e a atividade celular correspondente é predominantemnte excitadora, agrupando-se as descargas na fase ascendente e na descendente. No tipo com dois componentes (negativo e positivo) a atividade celular é caracterizada sobretudo por PIPS precedido de ativação. A evolução temporal do PIPS corresponde aproximadamente ao potencial positivo de superfície. A simplicidade dessas relações, contudo, é aparente, como se pode depreender das relações encontradas quando os potenciais são precedidos de um ligeiro pré-potencial. Tomando-se o ápice deste como zero vê-se que as descargas se agrupam em fase com êle e ocorre um PIPS em fase com o componente negativo do potencial de fuso (Fig. 3).

ORGANIZAÇÃO DA ATIVIDADE ELÉTRICA CEREBRAL
Os fatos acima descritos mostram que a manifestação eletrofisiológica resultante, que é a onda eletrocortical, pode envolver fenômenos causais diferentes. A manifestação de superfície da AEC em dado instante depende fundamentalmente do número de elementos ativados ou inibidos, das relações de fase temporal entre eles existente e do sentido das correntes originadas. Estas, obviamente, dependem da geometria de disposição dos neurônios, especialmente das interações dos seus prolongamentos. A complexidade destas últimas é impossível de ser totalmente decifrada. No córtex visual do gato, segundo Sholl18, cada fibra aferente entra em contato com cerca de 5.000 neurônios vizinhos. Isso significa que a ativação de uma fibra do trato genículo-calcarino pode alterar a excitabilidade de milhares de pontos sinápticos, o que se realiza por meio de PEPS e PIPS cujas correntes se compõem e contribuem para a AEC local. Embora a atividade de circuitos autóctones, em áreas restritas do córtex seja essencial para seu funcionamento e, conseqüentemente, para condicionar as oscilações da AEC, os impulsos que a eles chegam é que certamente os põem em atividade e estabelecem as interações intra e intercorticais e córtico-subcorticais. Burns4 desenvolveu técnica que permitiu estudar a atividade de pequenos blocos de córtex neuralmente isolados mas com sua circulação funcionante. Nessas condições verificou que o bloco isolado em geral não apresenta oscilações de potencial. Deixando, porém, pequena ponte de córtex de menos de 5 milímetros de largura entre o bloco isolado e áreas vizinhas já era suficiente para que oscilações reiteradas ocorressem, mostrando que a chegada de informações de outras áreas é essencial para que os circuitos locais entrem em ação. Esse fato é forte evidência experimental de que a atividade funcional de uma área cortical depende de sua interação com outras regiões corticais e subcorticais, uma vez que a atividade elétrica é a manifestação da atividade funcional.
Se os potenciais eletrotônicos e os potenciais de ação celulares atingem a superfície com amplitude suficiente para serem registrados, até mesmo através do couro cabeludo, é óbvio que há somação de múltiplos potenciais a cada instante. Quando se estuda a correlação entre os dois fenômenos eletrofisiológicos só é possível fazer o registro de uma ou de poucas células corticais. Disso resulta, como já dissemos acima, grande dificuldade para se estabelecerem com precisão as relações buscadas. Algumas tentativas, feitas por Verzeano e Negishi21, no sentido de se registrar a atividade elétrica de diversos neurônios tridimensionalmente, revelaram que mesmo em um espaço muito diminuto há uma seqüência definida de descarga em elementos distintos. Essa seqüência pode ser reproduzida quando se aplicam estímulos aferentes à área em estudo. Esses autores demonstraram que a ocorrência de ondas sincronizadas em um campo neuronal, registradas por macro-elétrodos, coincide com aumento da freqüência das espículas geradas pelos neurônios individuais e aumento do agrupamento periódico de tais potenciais.
Estabelecido que os potenciais do EEG correspondem à composição de PEPS, PIPS, potenciais de ação e oscilações muito lentas causadas por agregados neuronais corticais, a pergunta que surge em seguida é: por que razão alguns ritmos cerebrais ocorrem com freqüência baixa e com uma regularidade que surpreende, quando se considera o número de elementos corticais em atividade? Há diversas explicações possíveis para esse fenômeno: 1) a oscilação a freqüências regulares é fortuita e deve-se à simples composição de PEPS, PIPS, potenciais de ação e potenciais lentos que por acaso se compõem a tais freqüências; 2) há microcircuitos cerebrais que oscilam primariamente, devido a uma seqüência definida de PEPS, PIPS e potenciais de ação, e da composição ao acaso dessas oscilações primárias resultam os ritmos regulares; 3) há circuitos marca-passos que oscilam deterministicamente em certas freqüências e as impõem a outros circuitos corticais, originando harmônicos e/ou subarmônicos da AEC. O mais provável é que os três mecanismos desempenhem algum papel mas as provas disso ainda são fragmentárias e não permitem estabelecer com certeza a contribuição de cada um.
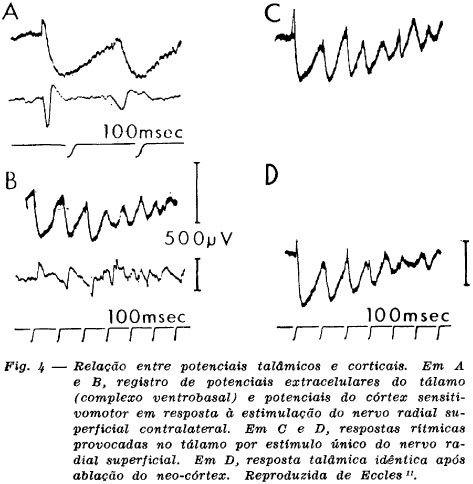
Eccles 9,10,11 demonstrou que há circuitos talâmicos que tendem a oscilar quando ativados adequadamente. Circuitos análogos podem existir também no córtex cerebral e no sistema límbico. A aplicação de um pulso aferente (estimulação de um nervo sensorial) provoca no complexo ventrobasal do tálamo (conjunto dos núcleos VPM e VPL) ondulação rítmica com descargas superpostas que ocorrem a intervalos sucessivos de 100 milissegundos. A estimulação do córtex somestésico causa na mesma região talâmica potenciais semelhantes. O registro intracelular de neurônios do complexo ventrobasal mostra PIPS de 100-200 milisegundos, seguidos de PEPS e espículas no ápice destes últimos. O registro simultâneo da AEC cortical mostra um potencial evocado por estimulação aferente, seguido de oscilações mais ou menos em fase com as alterações eletrotônicas registradas no complexo ventrobasal. A ablação crônica do córtex não abole as oscilações talâmicas, o que até certo ponto invalida a antiga teoria de que as oscilações que se seguem ao potencial evocado cortical demandam reverberação tálamo-córtico-tálamo-cortical (Fig. 4).
Os fenômenos descritos por Eccles podem ser explicados de acordo com o seguinte esquema: os neurônios do complexo ventrobasal enviam seus axônios ao córtex e recebem axônios do lemnisco medial; colaterais dos axônios eferentes ativam interneurônios inibidores que inervam recorrentemente os neurônios relés. Assim, a informação que é gerada nesses neurônios acaba gerando sua própria inibição recorrente por PIPS. Ao final dos PIPS os neurônios relés estão em fase de hiperexcitabilidade pós-anódica, a qual provoca PEPS. Sob a ação de interneurônios excitadores, que também estavam sob inibição e entram em hiperexcitabilidade pós-anódica, os neurônios relés podem descarregar impulsos; estes provocam ativação dos interneurônios inibidores, repetindo o ciclo. Uma vez que a extensão da inibição dos neurônios relés seja suficientemente grande esse mecanismo, por si só, pode ser responsável pela oscilação (Fig. 5).

A estimulação de núcleos mediais do tálamo provoca respostas de recrutamento cortical desde que a freqüência de estímulos seja baixa; a excitação a alta freqüência provoca dessincronização cortical. O recrutamento assim obtido é semelhante aos potenciais de fuso encontrados durante as primeiras fases de sono. Registrando-se os potenciais intracelulares de neurônios de núcleos ventrais do tálamo os estímulos de baixa freqüência aplicados aos núcleos mediais causam, também, PIPS; sua latência é grande (15-40 ms), o que sugere a existência de cadeias polissinápticas envolvidas na inibição. Com freqüências altas ocorre, porém, despolarização prolongada dos referidos neurônios, evidenciando a atividade de sinapses excitadoras. É importante lembrar que nessas circunstâncias o registro da AEC mostra dessincronização.
As ondas teta, de freqüência na faixa de 4-8 Hz, são características da AEC do hipocampo e outras estruturas límbicas no estado de vigília e na fase paradoxal do sono. Fujita e Sato13 mostraram que há relação entre as ondas teta e os potenciais intracelulares de células piramidais do hipocampo. Cada PEPS com descarga corresponde a urna onda positiva na superficie. Como Eccles havia demonstrado para o complexo ventrobasal, no hipocampo também há poderosa inibição recorrente, que entra em ação quando as células piramidais são estimuladas. A duração dos PIPS registrados é de 200-500 ms. Uma oscilação de potenciais com essa duração realiza-se, como se pode calcular, a freqüências muito baixas. Após o PIPS há um rebote excitador e as células piramidais emitem descargas (Fig. 6).

Os ritmos cerebrais regulares mais característicos, que são o ritmo alfa, o teta límbico e os fusos do sono, talvez sejam causados mediante combinação de fenômenos como os acima mencionados. Estes foram estudados, em geral, sob condições de estimulação elétrica transitória e, por isso, os referidos fenômenos também se apresentam com caráter efêmero. Em condições naturais, quando os ritmos alfa, teta e os fusos de sono ocorrem reiteradamente, deve haver mecanismos que os mantêm. Para o caso dos fusos do sono tais mecanismos são devidos a centros situados em núcleos da linha média e laminares do tálamo e no núcleo caudado. A obtenção de ondulação exatamente igual por estimulação das referidas estruturas é muito sugestiva de que essa localização seja verdadeira. No caso do ritmo teta hipo-campal pelo menos alguns desses circuitos situam-se na área septal. No que diz respeito ao ritmo alfa, porém, a situação dos marca-passos é totalmente desconhecida. Uma das razões disso é que não existe nos animais abaixo do homem modalidade eletrofisiológica que corresponda exatamente ao ritmo alfa, o que impede investigação técnicamente adequada. Os fusos registrados nas áreas frontoparietais dos mamíferos comumente utilizados em laboratorio são tidos, por alguns, como análogos ao ritmo alfa, porém essa afirmação não é correta, uma vez que tanto a morfologia dos potenciais como sua projeção cortical são inteiramente diversas. É certo, contudo, que o ritmo alfa é provocado pela atividade de algum sistema subcortical de projeção bi-hemisférica. Só assim se torna possível explicar a sincronia tão estreita, porém não absoluta, entre os potenciais alfa de ambos os lados. O desenvolvimento das técnicas de correlação 17 permitiu estudo muito acurado das relações de fase dos potenciais. Utilizando essa técnica Liske e col.16 estudaram as relações de fase entre as derivações P3-O1 e P4-O2, concluindo que não existe sincronia absoluta, sendo, porém, a diferença muito pequena; de 42 indivíduos normais examinados, em 24 encontraram precedência de fase no hemisfério cerebral direito (em média de 0,83 milisegundos).
No que tange aos potenciais de natureza patológica, como as descargas convulsivas e as ondas lentas decorrentes de edema cerebral, pouco se sabe sobre como são provocados. A presença de fenômenos inibidores recorrentes, de distribuição tão ampla e de intensidade tão elevada, deve ser importante para impedir que os circuitos corticais entrem em atividade, quer desordenada, quer sincronizada, provocando convulsões. A injeção de metrazol provoca ondas de superfície negativas simultâneas com despolarização celular8. De início esta droga ativa o córtex cerebral via sistema reticular ascendente19, provavelmente interferindo nas conexões dos dendritos apicais5, e depois provoca potenciais bifásicos (positivo-negativos) de grande amplitude. Nesta fase, que se deve à excitação direta do córtex cerebral, há despolarização maciça dos neurônios com potenciais de ação de alta freqüência agrupados 7.
Pelo que acaba de ser exposto conclui-se que o estudo da atividade elétrica cerebral, no que tange aos seus mecanismos, ainda esbarra com sérias dificuldades. Se a origem eletrofisiológica dos potenciais já começa a ser deslindada, graças ao desenvolvimento das técnicas de registro unitário do córtex cerebral e de estruturas subcorticais, os mecanismos operacionais de controle, que modulam as alterações do potencial de membrana em determinados neurônios, são ainda muito mal conhecidos. O prosseguimento dos estudos com micrométodos deverá trazer nos próximos anos dados significativos para a compreensão clara da origem e da função fisiológica das ondas cerebrais.
REFERÊNCIAS
1. ADEY, W. R. — Slow electrical phenomena in the central nervous system. Neurosci. Res. Progr. Bull. 7:77, 1969.
2. ANOKHIN, P. K. — The electroencephalogram as a result of ascending influences on the cells of the cortex. Electroenceph. clin. Neurophisiol. 16:27, 1964.
3. BRAZIER, M. A. B. — The historical development of neurophysiology. In Handbook of Physiology. Neurophysiology, vol. I, American Physiological Society, Washington, 1965, pp. 1-58.
4. BURNS, B. D. — The Mamalian Cerebral Cortex. E. Arnold, London, 1958.
5. COVIAN, M. R.; TIMO-IARIA, C. & MARSEILLAN, R. F. — Changes in cortical evoked potentials by previous reticular stimulation. In Brain Mechanisms and Learning. Blackwell Scientific Publications, Oxford, 1961, p. 443.
6. CREUTZFELDT, O. D.; FUSTER, J. M.; LUZ, H. D. & NASCIMENTO, A. — Experimenteller Nachweis von Beziehungen zwischen EEG — Wellen und Aktivität corticaler Nervenzellen. Naturwissenschaften 51:166, 1964.
7. CREUTZFELDT, O. D.; WATANABE, S. & LUZ, H. D. — Relations betwenn EEG phenomena and potentials of single cortical cells: evoked responses after thalamic and epicortical stimulation. Electroenceph. clin. Neurophysiol. 20:1, 1966.
8. CREUTZFELDT, O. D.; WATANABE, S. & LUZ, H. D. — Relations between EEG phenomena and potentials of single cortical cells: spontaneous and convulsoid activity. Electroenceph. clin. Neurophysiol. 20:19, 1966.
9. ECCLES, J. C. — The Physiology of Nerve Cells. Johns Hopkins,. Univ. Press, Baltimore. 1957.
10. ECCLES, J. C. — The Physiology of Synapses. Academic Press, New York, 1964.
11. ECCLES, J. C. — Inhibition in thalamic and cortical neurons and its role in phasing neuronal discharges. Epilepsia 6:89, 1965.
12. FROMM, G. H. & BOND, H. W. — The relationship between neuron activity and cortical steady potentials. Eletroenceph. clin. Neurophysiol. 22:159, 1967.
13. FUJITA, Y. & SATO, T. — Intracellular records from hippocampal pyramidal cells in rabbit during theta-rhythm activity. J. Neurophysiol. 27:1011, 1964.
14. GRUNDFEST, H. — The interpretation of electrocortical potentials. Ann. N. Y. Acad. Sci. 92:887, 1961.
15. LI, C. L. — Cortical intracellular synaptic potentials in response to thalamic stimulation. J. Cell. Comp. Physiol. 61:165, 1963.
16. LISKE, E.; HUGHES, H. M. & STOWE, D. E. — Cross-correlation of human alpha activity: normative data. Electroenceph. clin. Neurophysiol. 22:429, 1967.
17. SATO, K. — An interpretation concerning physiological significance of statistical nature of electroencephalogram. Fol. Psychiatr. Neurol. Jap. 10:284, 1957.
18. SHOLL, D. A. — The Organization of the Cerebral Cortex. Methuen & Co., London, 1956.
19. TIMO-IARIA, C. & WERNER, G. — Electroencephalographic changes caused by some analeptic drugs. Arch. Int. Pharmacodyn. 112:451, 1957.
20. UCHIDA, T. — Thalamo-cortical waxing and waning nature and activities of cortical neurons in the rabbit. Jap. J. Physiol. 16:389, 1966.
21. VERZEANO, M. & NEGISHI, K. — Neuronal activity in cortical and thalamic networks: a study with multiple mlcroelectrodes. J. Gen. Physiol. 43:177, 1960.
22. YAMAMOTO, C. & KAWAI, N. — Origin of the direct responses as studied in vitro in thin cortical sections. Experientia 23:821, 1967.
Departamento de Fisiologia — Faculdade de Medicina da Universidade de São Paulo — Caixa Postal 2921 — São Paulo, SP — Brasil.
Relatório apresentado ao II Congresso Brasileiro de Eletrencefalografia e Neurofisiologia Clínica (Porto Alegre, 5 a 9 de julho — 1970).





























